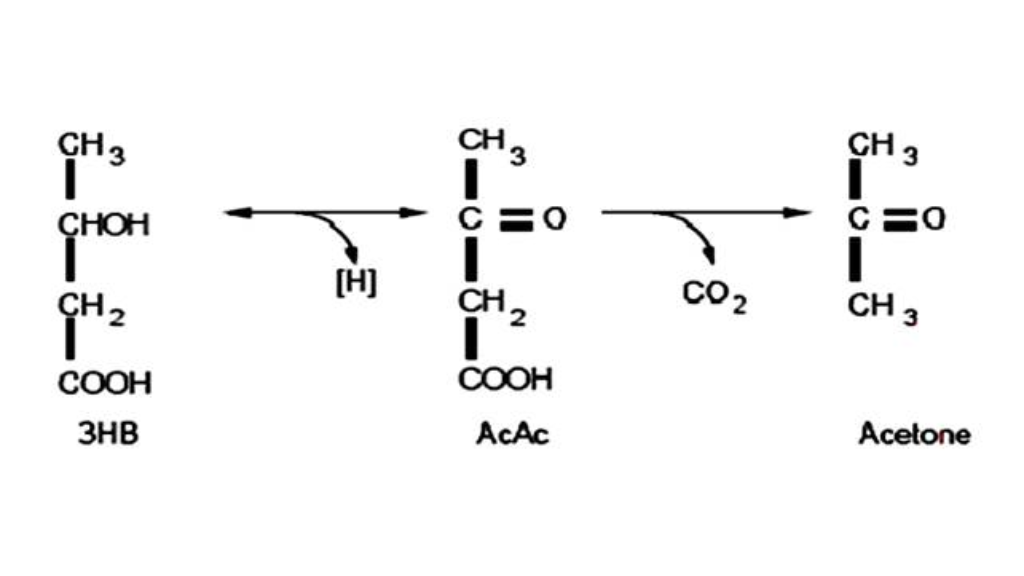
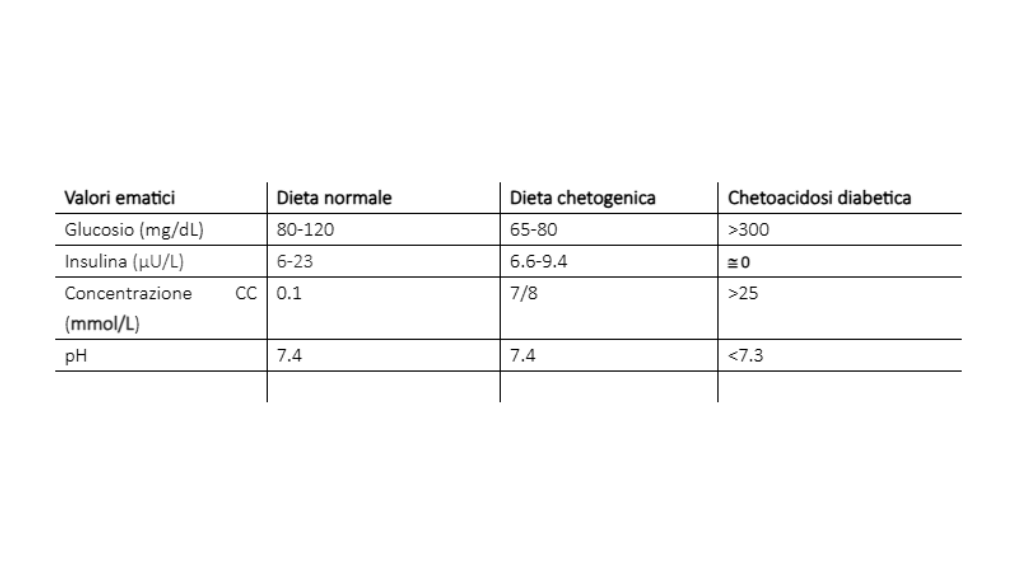
The term ketogenic diet is used to define a dietary regimen based on a drastic reduction in carbohydrate intake, associated or not with a relative increase in protein and fat intake (24). The metabolic state of ketogenic diets is relatable in many ways to fasting; in fact, even in fasting, that particular metabolic state known under the name of ketosis is established. The first in-depth scientific studies on this metabolic condition were those conducted by Cahill's group in the 1960s starting precisely from the "fasted" or "fasting" condition (9, 22). That of fasting is in fact a practice, or rather a technique, used for millennia to achieve particular states of spiritual well-being during rituals or religious practices. Even in the Old Testament as well as in the Qur'an and the Mahabharata there is mention of this ascetic practice.
We can find a reference to fasting, for example, in Matthew (17:14-21) where, in the episode of the 'healed epileptic, it is said, "This race of demons is not cast out except by prayer and fasting"; and it is no coincidence that fasting is mentioned in connection with epilepsy since it has been known since the 1920s that ketosis ( and thus fasting) is able to improve some types of epilepsy (29). Of course, one of the problems of fasting is the progressive depletion of the body's protein reserves. Modern ketogenic diets, on the other hand, seek to induce a state of ketosis while providing adequate protein intake so as to maintain lean mass; in fact, they have also often been called modified fasting "modified fasting diets" (5) or low carbohydrate and protein-sparing "low carbohydrate protein sparing modified diets" (3).